The Response of Predators to an Erupting Bison, Bison bison athabascae, Population
Nicholas C. Larter 1.3, Anthony R. E. Sinclair 1, and C. Cormack Gates 2
1 The Ecology Group
Department of Zoology
6270 University Boulevard
University of British Columbia
Vancouver, British Columbia
V6T IZ4
2 Department of Renewable Resources
P.O. Box 390
Fort Smith, Northwest Territories
XOE OPO
Current address:
Department of Renewable Resources
Bag Service # I
Inuvik, Northwest Territories
XOE OTO
Larter, Nicholas C., Anthony R. E. Sinclair, and C. Cormack Gates. 1994. The response of predators to an erupting Bison. Bison bison athabascae. population.
Summary
During the past 20 years Bison (Bison bison athabascae) numbers have increased and Moose (Alces alces) numbers appear to have decreased within the Mackenzie Bison Sanctuary. In adjacent peripheral areas near Mink Lake Moose densities were twice that in the Mackenzie Bison Sanctuary. Wolf (Canis lupus)activity was greater in the Mackenzie Bison Sanctuary than in the Mink Lake area. Although Bison made up a larger proportion of the Wolf diet (based upon scat analysis) in the Mackenzie Bison Sanctuary than in the Mink Lake area, Moose made up a significantly greater (P<o.oo1) proportion="" of="" the="" wolf="" diet="" than="" expected="" given="" availability="" prey="" biomass="" in="" both="" mackenzie="" bison="" sanctuary="" and="" mink="" lake="" areas.="" that="" moose="" made="" up="" a="" similar="" areas,="" there="" was="" two-fold="" difference="" densities="" between="" area,="" predation="" may="" be="" destabilizing="" exacerbating="" decline="" numbers.<="" p="">
Key Words: Bison, Bison bison athabascae, population eruption, Moose, Alces alces, Wolves, Canis lupus, Mackenzie Bison Sanctuary, Northwest Territories.
Reprinted from Canadian Field-Naturalist 108(3): 318-327.
The Response of Predators to an Erupting Bison, Bison bison athabascae,Population
Bison (Bison bison athabascae as defined by Van Zyll de long (1986) but see Geist 1992 for a contrasting view on the validity of subspecies in Bison) were reintroduced into the Mackenzie Bison Sanctuary in 1963. The population has increased rapidly (Gates and Larter 1990) and the current population eruption has followed the dynamics predicted by the Caughley-Riney four-stage equilibrium model (Riney 1964; Caughley 1970) despite the presence of Wolf(Canis lupus) predation (Larter 1994). This four-stage equilibrium model predicts that an erupting ungulate population will go through four stages; (1) a progressive increase in population size in response to the disparity between the number of animals present and the carrying capacity of the environment, (2) a levelling off in animal numbers in response to decreasing forage availability, (3) a decline in numbers because the population has increased beyond the carrying capacity of the environment, and (4) a phase of relative stability with population density remaining lower than peak density because the carrying capacity of the environment has been reduced by the impact of peak population density. As the population passes through the four stages, food supplies are reduced and the plant community becomes increasingly dominated by more unpalatable species in response to increased grazing intensity. Beyond documenting that this model's predictions appear to hold for an indigenous ungulate population in the presence of predators, further important questions arise regarding the response of the Wolf population to an increasing prey base. Do Wolf numbers increase in response to the increasing abundance of Bison? If Wolf numbers increased, what impact could this have on the Bison population, and on Moose (Alces alces), the only other ungulate prey population in this study area? Gates and Larter (1990) proposed that an increasing Bison population in the Mackenzie Bison Sanctuary and adjacent areas, may not be diverting Wolf predation from Moose and Caribou by providing a substantial alternate prey source; rather it could be increasing it by maintaining high Wolf populations capable of depleting the more vulnerable prey populations.
In systems where there is an alternate prey source and prey switching occurs, it is theoretically possible for predation rate to be either directly density-dependent or inversely density-dependent over all densities (Ricklefs 1979). Inverse density-dependent predation rates occur when predation rate increases while prey density decreases, a situation that could be destabilizing and potentially drive a prey population to extinction.
The vulnerability of the primary prey relative to an alternate prey determines the impact of predation on the abundance of primary prey (Messier 1993). Jarman and Johnson (1977) suggested that the decline in introduced hare (Lepus europaeus) and native rat kangaroo (Aepyprymnus spp.) population in Australia may have been a result of introduced fox (Vulpes vulpes) populations being sustained by rabbits (Oryctolagus cuniculus). Consequently foxes may have been able to eliminate a less numerous and more catchable prey. A similar situation was described in England where fox numbers were being maintained by rabbits at numbers high enough to significantly reduce breeding densities of Gray Partridge (Perdix perdix) (J. Reynolds, personal communication). Rock-wallaby (Petrogale lateralis) populations in Western Australia are being decimated by fox predation (Kinnear et al. 1988), because fox numbers are being kept high by a substantial rabbit population.
In some areas of western North America, Moose are the principal prey of Wolves which can remain at high enough population levels to deplete Caribou (Rangifer tarandus) populations. Seip (1992) documented that a Woodland Caribou (R, t. caribou) population separated from Wolves and Moose during summer, and therefore less vulnerable to Wolf predation, had a lower adult mortality rate and higher calf survival than a similar Woodland Caribou population that was not spatially separated from Wolves and Moose. He believed that because the Wolf population was sustained primarily by Moose, the latter Caribou population could potentially be extirpated.
We hypothesize that predators are capable of utilizing both Moose and Bison in this system, and that Wolf numbers have increased in response to increasing Bison numbers. we predict that over a range of Moose densities Wolves will continue to utilize Moose to a greater extent than expected given Moose and Bison availabilities thus providing the potential for local extinctions of Moose populations.
In this paper we examine Wolf-Bison-Moose dynamics in two areas by comparing Wolf diet, Wolf abundance, relative prey abundance and available prey biomass to test whether Wolf predation on Moose, in the presence of Bison, can act in a destabilizing and inversely density-dependent fashion. Because areas that have been proposed for future Bison reintroductions have low densities of Moose and Wolves, and a subsistence Moose harvest, knowledge of Wolf-Bison-Moose dynamics is becoming increasingly important.
Materials and Methods
Study Area
In 1963, eighteen Bison were released into an unpopulated area of their historic range on the western side of Great Slave Lake (61030' N, 117°00' W) in the Northwest Territories of Canada. The population increased in an eruptive fashion (Calef 1984), and Bison now inhabit an area exceeding 9000 km2 (Larter and Gates 1994). Recently, the population split into two distinct subpopulations: the Mackenzie subpopulation (MBS) inhabiting the core of the range and the Mackenzie Bison Sanctuary proper, and the Mink Lake subpopulation (ML) inhabiting the more peripheral areas of the range, the Mink Lake area northwest of the Sanctuary. The study area is located in the Upper Mackenzie Section of the Boreal Forest Region (Rowe 1972) in the emerged bed of a once vast glacial lake. The undulating topography is dominated by forested habitats which include coniferous forests of Black Spruce (Picea mariana), White Spruce (P. glauca),Jack Pine (Pinus banksiana), and Larch (Larix laricina), and mixed deciduous- coniferous forests of White Spruce, Trembling Aspen (Populus tremuloides), and Balsam Poplar (P. balsamifera). Shallow lakebeds are widely dispersed throughout the area. These lakebeds are gradually filling in with sedges and grasses, while woody plants, notably willows (Salix spp.), are invading these lakebeds and provide most of the forage used by Bison, but represent <6% of the study area (Larter and Gates 1991a; Matthews 1991).
Predator Abundance/Activity
We calculated indices of Wolf abundance in both areas (MBS and ML) using two techniques: track counts and visual observations. Track count lines were established during winter 1989-1990 and consisted of three, 1 km transects along seismographic survey lines in each area. The start and end points of each transect were flagged for identification and track counts were made on them throughout the winter when conditions permitted. The number of track lines was increased to five and the transect length increased to 3 km during winter 1991-1992 because of the number of zero counts during 1989- 1990. No data were collected during winter 1990- 1991. Data were collected during three winter periods: early (November), mid- (February), and late- winter (March-April). A track-day was defined as a 24-hour period when all new tracks along the trail were made within the preceding 24-hours. Because there were different numbers of successful track- days between areas, all data were expressed as number of tracks counted per track-day. Because of reduced sample size we combined track-day data from the two winters, and used a square root transformation to normalize the data. The Student's t-test was used to determine if there were differences- in the number of tracks/track-day between areas.
Wolves were observed during 25 hours of aerial reconnaissance, and ground-based travel throughout the study area. Abundance indices for both areas were calculated as the number of Wolves observed per day of field work, or the number of Wolves observed per flight-hour.
Prey Abundance and Availability
Population estimates for Bison were usually conducted on a biannual basis during 1963-1992. Prior the 1989 a total count technique was used. This was changed to a stratified strip transect technique ion 1989. Sex age composition surveys were conducted the yearly from 1985-1992 (Gates et al. 1991). Because of logistic problems with the 1992 census, we used the 1989 survey data to estimate numbers of animals in the different sex and age classes. We multiplied the numbers of each sex and age class by a winter weight estimate to determine available Bison biomass. Winter weight estimates for Bison were: calves 200 kg, yearlings 360 kg, adult females 590 kg, and adult males 850 kg (Larter and Gates 1990; Gates unpublished data).
Aerial transect surveys for Moose were conducted just in the Mackenzie Bison Sanctuary in February 1965 and December 1971 (Department are Renewable Resources, Government of the Northwest Territories unpublished data). The number of Moose are observed per kilometre of transect was determined. In April 1987 while conducting the Bison census of the Mackenzie Bison Sanctuary and Mink Lake area we also calculated the number of Moose observed per kilometre of transect. During November 199l a random stratified survey (Gasaway et al. 1986) was conducted to estimate Moose densities in both areas (Shank 1992).
Additional indices of abundance were collected, from aerial- and ground-based reconnaissance of study area by personnel during 1986 to 1992. The ground-based index was calculated as the number of Moose observed per day of field work.. For convenience, this was then converted into Moose observed per week of field work. The aerial based index was calculated as the number of Moose observed per flight-hour.
Available biomass of Moose was estimated as for Bison. We used Moose sex/age ratios and population estimates collected during the 1991 census (Shank 1992). Weight estimates for Moose were: calves 160 kg, yearlings 240 kg, adult females 350 kg, and adult males 453 kg (Blood et al. 1967; Ritcey 1974; Banfield 1977; Saether 1985).
Wolf Scat Analysis
Wolf scats were collected opportunistically throughout the course of the study. Scats found near Wolf kills were not collected because they could test bias the data set. Scats collected in winter were kept in frozen, while scats collected in summer were dried prior to preparation for laboratory analysis followed Kennedy and Carbyn (1981). Bone fragments, hair, and feather characteristics were used to determine the prey item(s) present in scats.
We compared the presence of prey items in Wolf scats between study areas during winter and between seasons, for the Mackenzie Bison Sanctuary (MBS) area only, using the proportion test (Zar 1984 ). Small sample size precluded comparing seasonal differences for the Mink Lake (ML) area. We calculated available prey biomass (Bison and Moose) for each area (see Prey Abundance and Availability). We used the log-likelihood ratio (G-test, Zar 1984) to compare the frequency of scats containing the different prey items with that available (live biomass ) in each area.
We estimated relative amounts of Bison and Moose consumed following the relationship described by Floyd et al. (1978).
y = 0.38 + 0.02x
where: y = kg of prey/collectible scat
x = the average weight of individuals of each prey type.
We estimated x for each prey in each area by determining the proportion of juveniles and adults killed based on Wolf kill data, and multiplying that proportion by the average weight of juveniles and adults. Juvenile weights are an average of calf and yearling weights and adult weights are an average of male and female weights. We calculated y for each prey, and multiplied y by the number of scats in which the prey items occurred. We multiplied by frequency of scats because the majority of scats (77%) contained only one major prey item (either Bison or Moose). This calculation was done for each area and the relative amounts of each prey species was expressed as a ratio of kg of Bison consumed per kg of Moose.
Wolf Kills
\Wolf kills were located opportunistically by ground-based and aerial reconnaissance. Whenever possible, kills located from the air were verified by ground observations. Evidence that an animal was killed by Wolves includedobvious chase sequences in the snow and/or signs of struggle before death (i.e. extensive blood covered vegetation or snow, and broken vegetation). The sex/age class and species of prey were determined by physical examination.
Results
Predator Abundance/Activity
Overall, Wolf activity was greater in the Mackenzie Bison Sanctuary (MBS) than in the Mink Lake (ML) area during all three periods of the winter (Table 1). Differences between areas were not significant when compared at anyone sampling period (P = 0.063, t = 1.58, early winter; p = 0.65, t = - 0.40, mid-winter; p = 0.37, t = 0.33, late winter). However, when the data were lumped across the entire winter the difference became significant: 0.43 and 0.21 mean tracks/track-day in MBS and ML respectively (P = 0.021, t = 2.07).
TABLE I. Comparative Wolf activity (tracks/track-day) between the Mackenzie Bison Sanctuary and Mink Lake area during three winter periods: early (November), mid (February), and late (March-April). The number in parentheses is the number of track days used to calculate tracks/track-day. ** indicates significance between areas at p < 0.05, * indicates significance at p
|
Winter |
||||
| Area | Early | Mid | Late | Lumped |
| MBS | 0.65 (17)* | 0.09 (11) | 0.43 (37) | 0.43 (65)** |
| ML | 0.23 (26)* | 0.14 (30) | 0.27 (22) | 0.21 (78)** |
Ground-based and aerial observations showed a similar pattern to the track count data, but with greater disparity between areas. The aerial-based index of abundance was twice as high in MBS than ML averaging 2.2 Wolves observed/flight hour (n= 16.5 flight hours) versus 1.2 (n=8.5 flight hours), respectively. The ground-based index was four times as high in MBS than ML averaging.0.31 Wolves observed/field day (n = 571 field days) versus 0.07 (n = 133 field days) respectively.
Prey Abundance
There were approximately three times as many Bison in MBS as at ML (1885 and 546 respectively). When these data were converted from individuals observed to number of groups (or search items) counted, there were 138 Bison versus 79 Moose groups and 48 Bison versus 190 Moose groups in the MBS and at ML respectively. The estimated available biomass of Bison per total area was 1106 tonnes for MBS (4563 km2 area) and 343 tonnes for ML (3500 km2 area) (Figure 2A).
The 1965 and 1971 transect surveys for Moose in the MBS gave similar results: 87 Moose along 1834 km of transect, and 82 Moose along 1496 km of transect respectively. In 1987, one Moose was observed in ML on 1075 km of transect covering both the MBS and ML. Moose densities in 1992 were 0.12 and 0.25/km2in MBS and ML respectively (Shank 1992). Abundance indices based upon ground and aerial reconnaissance data from 1986-1992 also indicated lower Moose numbers in MBS relative to ML: 0.21 Moose observed/field week (n =46.7 field weeks) versus 0.63 (n = 14.3 field weeks) respectively, and 0.9 Moose observed/flight hour (n = 30.9 flight hours) versus 1.6 (n = 11.5 flight hours) respectively. The estimated available Moose biomass was 46 tonnes for MBS (4563 km2 area) and 91 tonnes for ML (3500 km2 area) (Figure 2A).
Predator DietAlthough Bison and Moose made up a large proportion of the diet, small mammals including mice (Cricetidae), Snowshoe Hare (Lepus americanus), and birds (both Gallinaceous and waterfowl) were also consumed. The number of scats containing Bison and Moose was equal in ML during winter. In MBS, Bison was present in more scats than Moose (Figure 3). The proportion of scats from MBS containing Moose and Bison was similar (P>O.2) between sea- sons (Z = 0.93 for Moose, Z = 0.09 for Bison, snow free=summer, snow cover=winter). During winter the proportion of scats containing Bison was greater in MBS than that at ML (P<o.05, <="" em="">Z = 2.91); there was no difference in the proportion of scats containing Moose (P>0.3, Z = -0.58). Insufficient sample size prevented the seasonal comparison of scats found in ML. The number of scats containing Moose was significantly greater than that expected given the available biomass of Moose and Bison in both ML and MBS (G = 340.0 ML, G = 1624.3 MBS,P<0.001) (Figure 3). Wolves consumed 1.62 kg of Bison per 1 kg of Moose in MBS, compared to 1.17 kg of Bison per 1 kg of Moose in ML.
Forty-one of 46 documented Wolf kills were Bison. Of the 41 Bison kills, 31 were juveniles: 26 calves and 5 yearlings (Figure 4). The greater proportion of Bison, especially juveniles, in Wolf kills was more pronounced in MBS. Moose made up a larger proportion of kills at ML than at MBS (Figure 4).
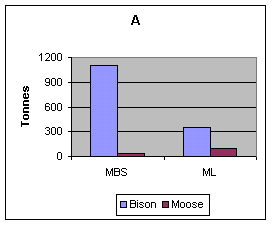
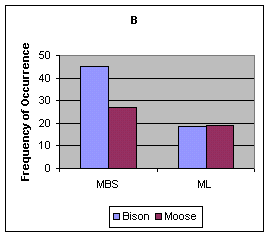
FIGURE 2. (A) Available Bison and Moose biomass in the Mackenzie Bison Sanctuary and Mink Lake area, (B) frequency of occurrence of Bison and Moose in Wolf scats (during winter) in the Mackenzie Bison Sanctuary and Mink Lake area. n = total number of scats.



Prey Type
Figure 3. Percent frequency occurrence of various prey items found in Wolf scats during different seasons and in different areas. Values above histograms are the number of occurrences. n=number of scats.
Discussion
Whether Moose numbers have declined over th entire study area from 1971 to the present is debatable. The crude line transect surveys do not provide estimates of Moose density, and any density estimates derived from them would be underestimate (Gasaway et al. 1986). However, local hunters maintain that Moose numbers were historically much higher in the MBS in the early 1970s, and the actual numbers observed during line transect surveys in the MBS do show a decline. Currently Moose number and density are lower in MBS than ML, and likely have been since 1986. Bison numbers have been increasing since the 1963 reintroduction. The increase has most likely had a greater potential impact on Moose in MBS than ML based upon total numbers of resident Bison and the length of time Bison have been resident in the two areas. Decreased Moose densities could have resulted from completion between Moose and Bison for food resource changes in habitat, or Wolf predation.
Competition for food resources is unlikely. Moose are concentrate selectors (Hofmann 1989) with die dominated by browse species like willow, aspen, and Paper Birch (Betula papyrifera) (Belovsky 197: Peek 1974; Risenhoover 1989). In contrast, Bison are classic grass/roughage eaters (Hofmann 1984 with diets dominated by graminoids (Reynolds et al. 1978; Larter and Gates 1991a, 199Ib). During fall (September-October) Bison in the MSB also utilize, lichen (Larter and Gates 1991a, 199Ib). In exceptionally dry summers, when grass and sedge productivity is reduced, Bison have foraged on willow (Larter and Gates 199Ia), but they rarely foraged willow during winter. Consequently, any dietary overlap would be limited in duration and occur during the growing season when forage quality and quantity are highest.
Habitat changes resulting in decreased willow cover could cause declines in Moose numbers. Aerial photos indicate that over the past 30 years, willows have invaded dry meadow communities and willow cover has actually increased, although some of the willow cover may have grown out of reach for Moose browsing. Fires have occurred more recently in ML than MBS, but have been infrequent during the past 30 years. It is unlikely that habitat changes causing a reduction in browse availability are responsible for declining Moose numbers. Predation remains an alternate possibility.
Wolves have historically been present in the area (Wood Bison Recovery Team 1988; A. Look, personal communication). Local residents believe that Wolf numbers are on the rise, but unfortunately there are no data on Wolf numbers during most of this time period. Messier and Crete (1985) have provided the best evidence of Wolves regulating their prey population. They documented Wolf predation over a range of Moose densities, and concluded that
Wolf predation acted in a density-dependent fashion. They found at low densities of 0.22 and 0.17 Moose/km2, Wolves were capable of regulating Moose populations. However, at densities around 0.40 Moose/km2, Moose numbers appear to be regulated by food. On Isle Royale, Moose numbers were regulated by food even in the presence of Wolves, and Moose densities have fluctuated between 1.6 to 2.8 Moose/km2 (from Peterson 1977, 1992). Messier (1991) estimated that competition for food had a regulatory effect on Moose density on Isle Royale when densities were between 1-2 Moose/km2. Crête (1989) estimated that densities greater than 2.0 Moose/km2 were required if carrying capacity (K) (Macnab 1985) was to be reached in eastern Quebec. Messier (1984) suggested that a density of 0.2 Moose/km2 approximated a threshold below which Wolf packs cannot subsist without an alternate ungulate prey species. Current Moose densities in the MBS are 0.12 Moose/km2 (Shank 1992); i.e., below Messier's (1984, 1993) threshold.
Wolf predation on Moose can represent a destabilizing influence which can potentially exacerbate a Moose decline because of its antiregulatory effect (Messier 1991). Additionally, Wolf predation has a greater impact on Moose populations when the Moose population is declining (Gasaway et al. 1983). Our data support the contention that Wolf predation on Moose is destabilizing, because in areas of both low and high Moose density, predation on Moose is greater than expected given the available prey biomass.
Moose remains were found in significantly more Wolf scats than would be expected given the availability of Moose and Bison biomass in the study area (Figure 2). This was not the case for Wolf kills. The proportions of Bison to Moose kills (Figure 4) were similar to that expected given prey availability (Figure 2A). The difference between data sets may be an artefact of sampling methodology. The majority of ground-based travel was through open habitat patches and along travel corridors that connect these open habitat patches. Consequently. travel was concentrated in Bison habitats as opposed to Moose habitat. Wolf kills were generally found by ground-based travel and were much easier to spot in the more open habitats where travel was concentrated. Sightability of animals and kills in forested habitats is restricted (Larter 1988).
Wolf scats were found opportunistically, were not collected from recent kill sites, and likely represent a less biased sample of Wolf diet composition than kill data. Estimates, derived from wolf scats, of the relative amounts of each prey item consumed were 1.62 kg Bison per 1 kg Moose in MBS and 1.17kg Bison per 1 kg Moose in ML. Given that the available biomass of Bison is 23.8 times that of Moose in MBS and 3.8 times that in ML, these data further indicate Moose as the preferred prey item.
Bison and Moose are the two largest species of North American ungulates. The typical attack success of Wolves ranges from 5% on larger prey (Haber 1977) to 40% on smaller ungulates (Kolenosky 1972). Given a choice, Wolves exhibit a preference for moderate size prey species (Murie 1944; Mech and Frenzel1971; Van Ballenberghe et al. 1975; Carbyn 1983). Only juvenile Moose and Bison are small, and the smaller size of Moose compared to Bison may affect prey preference.
The spatial distribution of prey can alter the effective search of Wolves and, regardless of prey preference, may affect prey selectivity (Huggard 1991 ; 1993). Clumping of prey means a lower effective search because the group, not the individual, represents a search item (Taylor 1979; Huggard 1991). Bison are gregarious, spending much of the year in large groups dominated by females, calves, and immature animals (Larter 1988). Moose are relatively solitary animals (Geist 1963; Banfield 1977). Consequently even though available prey biomass is skewed highly toward Bison, the actual number of search items may be closer to parity. When our census data were converted from individuals to the number of search items this was the case. However, there is some evidence that hunting success is decreased for predators that stalk groups rather than solitary prey (Van Orsdol 1984), and Moose tend to be well dispersed, spatially predictable, and always part of a year-long prey base (Messier 1993) therefore, Moose may be a more vulnerable and selected prey than Bison regardless of the number of search items.
Selectivity by the predator, especially in regard to the sex/age classes of the different species which they prefer to attack, can shape the prey populations in different ways (Huggard 1991; Mills and Shenk1992). Consequently predation may affect Bison and Moose populations quite differently. Our data are limited, but the majority of Bison killed by Wolves were juveniles, whereas the majority of Moose killed were adults. If adults are taken in a greater proportion than juveniles, predation will have a greater impact on the prey population because it directly influences fecundity and mortality.
Wolves do not remain in areas of high prey density, they travel frequently throughout their territory
(Carbyn 1983). Wolves preying upon Bison in Wood Buffalo National Park (WBNP) have a long average distance between kills suggesting that they must travel frequently between herds for kills. Although previously attacked Bison herds are more alert Wolves will occasionally follow these herds (Oosenbrug and Carbyn 1985). Wolves may be more successful when they encounter large herds because in of the higher probability of finding weak or vulnerable individuals, however vulnerability does not necessarily imply weaker or sick individuals.
Wolves attack Bison herds with calves preferentially over herds without calves (Carbyn and Trottier1987). This preference for juveniles and especially calves would be expected because calves are generally bit a slower, less dangerous and more inexperienced with predators. Calves are easier to kill than adults (Carbyn and Trot tier 1988), however, adult Bison and are still taken. Van Camp (1987) found that in the Slave River Lowlands (SRL) adjacent to WBNP five packs of Wolves killed and consumed 15 adult (13 females and 2 males) and four calves duriing a nine-week period in late winter.
Calf remains in scats are indistinguishable from other age classes after three months because calves lose their red pelage. Therefore any scats collected in winter provide no information upon whether the prey was juvenile or adult. However, 26 of 41 Bison killed by Wolves were calves. Radio-collared Bison and sequential cow:calf ratios from the study area (Larter 1994) indicate that Bison calves are preyed upon year-round, but predation pressure may increase during the summer on newborn individuals. Sequential adult female:calf ratios during summer 1991 in the MBS show a decline between June and August: 46.7 + 6.7 to 36.8 +11.7 calves: 100 adult females. During summer radio-collared calves were lost to Wolf predation at a rate of 1 calf/153 collar-days, but during winter the rate decreased to I calf/354 collar-days (Larter, unpublished data). The possible preference for more vulnerable newborn and Bison calves as prey during summer may have important consequences for juvenile Wolf survival. It is during summer that pups are being reared and foraging radius is reduced.
Hunting success of Wolves generally increases in deep snow (Kolenosky 1972; Haber 1977) because deep snow hinders ungulate movements. Snow depths of 50-55 cm hinder Bison calves (Van Camp 1975), whereas depths of 80-84 cm are required to hinder Moose calves (Kelsall 1969). Adult Bison can still successfully forage at snow depths of 75-85 cm and snow densities of up to 0.2 g/cm3 (Van Camp 1975), while adult Moose movements become hindered at 98-105 cm (Kelsall 1969). Snow levels in the study area commonly reach 50 cm by mid-winter (Larter 1988; Larter and Gates 1991a), but rarely reach 80 cm. The greatest depth recorded was 82 cm during late winter 1991. Therefore, Bison calves should be the most susceptible prey item during winter. Bison calves made up a larger proportion of Wolf kills than any other prey class (species, sex, and age).
If the Wolf population has been increasing in response to an increasing alternate prey base, then the probability of a random encounter with Moose also increases, Travel between open habitat patches (Bison habitat) necessitates travel through forested Moose habitat. If a pack of Wolves encountered Moose while actively searching for Bison it is unlikely that they would pass up the opportunity to attempt a kill, especially when the likelihood of success is higher.
In order to critically evaluate whether Wolf predation on Moose is acting in an inverse density-dependent manner it would be necessary to have a measure of Wolf numbers (i.e., the numerical response), and a measure of killing rate per predator (i.e., the functional response). Our data do not provide these definitive measurements, however they do provide a comparison of Wolf activity, prey abundance, and Wolf diet composition between two areas. The index of Wolf abundance (Wolf activity) is different between areas and consequently can be considered as an estimate of the numerical response of Wolves to an increasing Bison population over time. The Wolf diet composition in conjunction with prey (both Moose and Bison) abundance could be thought of as providing a crude estimate of the functional response. Since the frequency of Moose remains occurring in Wolf scats is similar between areas, while the availability of Moose is lower in the high Wolf density area, Wolf predation on Moose is potentially increasing as the Moose population declines. This situation could produce an inverse density-dependent relationship.